微丝
微丝(microfilament,MF)又称肌动蛋白丝(actin filament),是由肌动蛋白(actin)组成的细丝。它以束状、网状或散在等多种方式有序地存在于细胞质的特定空间位置上。
和微管一样,微丝是不稳定的,但它在细胞中也能形成如肌肉细胞中的收缩单位一样稳定的结构。微丝与许多种肌动蛋白结合蛋白相结合。
一、肌动蛋白与微丝的结构
微丝的主要成分是肌动蛋白,与微管相比,肌动蛋白微丝更纤细柔顺。
单个微丝通常比微管短很多,在细胞内单条微丝并不是独立行动的,而是形成横向连接的聚合物或形成束。
(一)肌动蛋白的结构
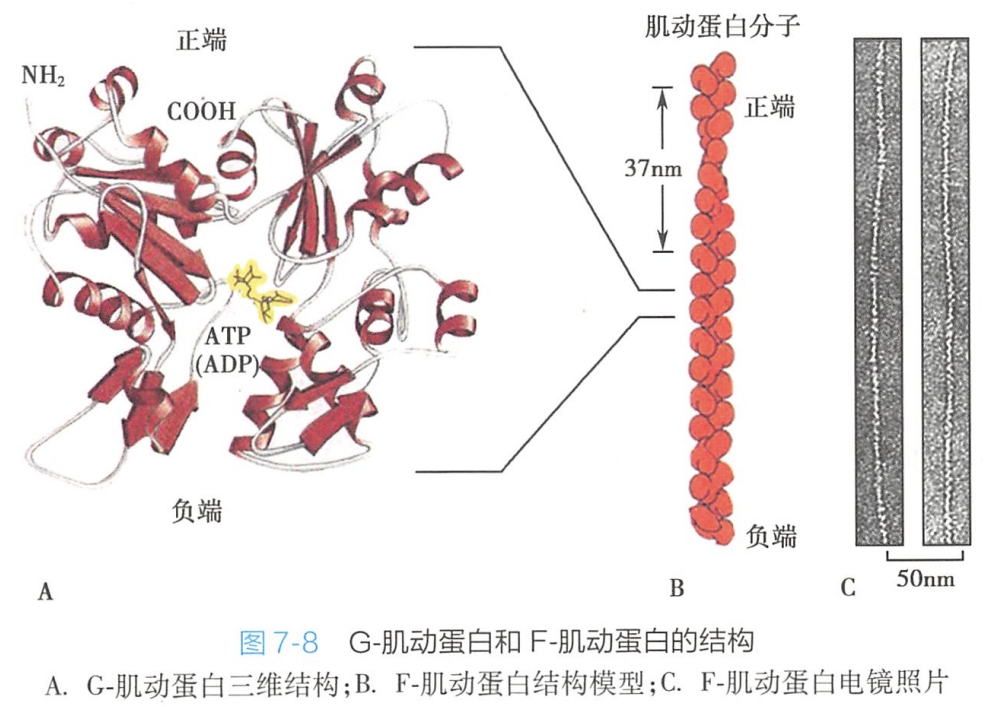
每一个肌动蛋白分子是由375个氨基酸组成的单链多肽,与一分子ATP紧密相连。肌动蛋白单体外观呈哑铃形,称为G-肌动蛋白(球形-肌动蛋白)。每个G-肌动蛋白由两个亚基组成( #? 单链怎么有两个亚基呢哥哥),它具有阳离子(Mg2+和K+或Na+)、ATP(或ADP)和肌球蛋白结合位点。
(二)微丝的结构
微丝是由G-肌动蛋白单体形成的多聚体,也称为F-肌动蛋白(纤维状-肌动蛋白)。肌动蛋白单体具有极性,装配时首尾相接,故微丝也有极性。肌动蛋白微丝也和微管一样是极性结构,有两个在结构上不相同的末端,相对迟钝和生长慢的负端(minus end)及一个生长快的正端(plus end)(图7-8)。负端又称为“指向端”(point end),正端又称为“秃端”(barbed end)。
一个细胞内的微丝总长度比微管总长度长至少30倍以上,反映了这两种细胞骨架聚合体在组装及功能上的差别。
二、微丝结合蛋白及其功能
纯化的肌动蛋白在体外能够聚合形成肌动蛋白纤维,但是这种纤维不具有相互作用的能力,也不能行使某种功能。体外组装的肌动蛋白纤维本身同活体中的肌动蛋白纤维之间没有差别,只是组织结构不同。
体内的肌动蛋白纤维是由不同的肌动蛋白纤维结合蛋白将肌动蛋白纤维组织成各种不同的结构,从而执行不同的功能。肌细胞和非肌细胞中都有微丝结合蛋白,至少已分离出100多种。
三、微丝的装配机制
在多数非肌肉细胞中,微丝是一种动态结构。
(一)微丝的组装过程
体外实验表明,球形肌动蛋白组装成肌动蛋白纤维需要ATP和一定的盐浓度(主要是Mg2+和K+),其组装过程也分三个阶段:即成核期、聚合期和稳定期。
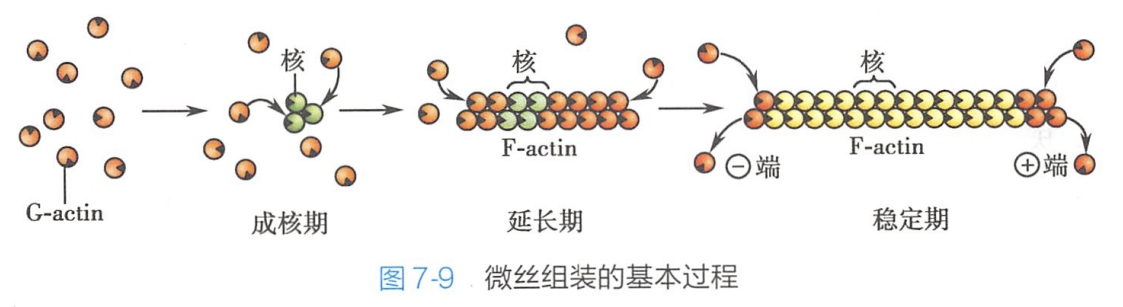
- 成核期是微丝组装的限速过程,故又称延迟期,此期球状肌动蛋白开始聚合,其二聚体不稳定,易水解,只有形成三聚体才稳定,即核心形成。
- 一旦核心形成,球状肌动蛋白便迅速地在核心两端聚合,进入聚合期。微丝两端的组装速度有差异,正端的组装速度明显快于负端,约为负端的10倍以上。
- 微丝延长到一定时期,肌动蛋白渗入微丝的速度与其从微丝上解离的速度达到平衡,此时即进入平衡期,微丝长度基本不变,正端延长长度等于负端缩短长度,并仍在进行着聚合与解聚活动(图7-9)。
(二)踏车模型和非稳态动力学模型
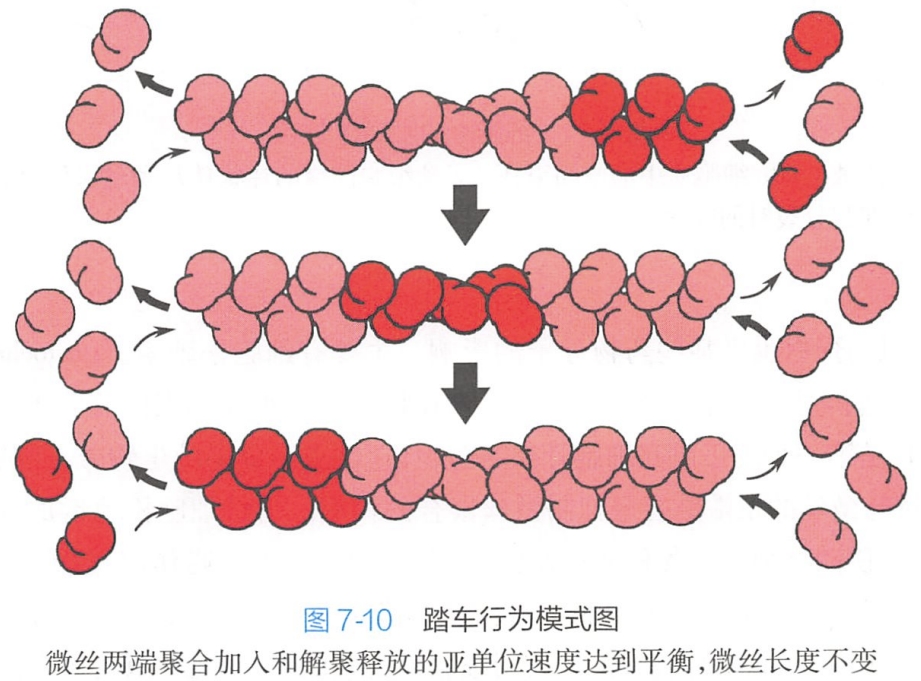
1.踏车模型
目前认为踏车模型在微丝组装过程中可能起主导作用。微丝可以在任何一端以添加肌动蛋白单体的方式增长,但由于微丝具有极性,新的肌动蛋白单体加到微丝两端的速度不同,速度快的一端为正端,速度慢的一端为负端,表现出显著的踏车现象。在微丝装配时,肌动蛋白分子添加到肌动蛋白丝上的速率正好等于肌动蛋白分子从肌动蛋白丝上解离的速率时,微丝净长度没有改变,这种过程称为肌动蛋白的踏车行为(图7-10)。
2.非稳态动力学模型
裸露的微丝就像不带结合蛋白的微管一样,呈固有的不稳定性,而且两端都可以解聚,在解聚时正端的速度比负端的速度要快得多。
非稳态动力学模型认为ATP是调节微丝组装的动力学不稳定性行为的主要因素。微丝组装的延长期通过ATP来调节,一个球状肌动蛋白分子可结合1分子ATP,结合ATP的肌动蛋白(即ATP-肌动蛋白)对纤维状肌动蛋白末端的亲和性高,当ATP-肌动蛋白结合到末端后,肌动蛋白的构象发生改变,ATP水解为ADP+Pi。ADP-肌动蛋白对纤维末端的亲和性低,容易从末端脱落,使纤维缩短。
ATP-肌动蛋白浓度与其聚合速度呈正比,当ATP-肌动蛋白高浓度时,ATP-肌动蛋白在末端聚合的速度便升高,由于ADP-肌动蛋白对末端的亲和力小,结果ADP-肌动蛋白不断从末端解聚脱落,使纤维缩短。
因此,就每一根纤维来说,其长度一般不是固定不变的,而是呈动力学不稳定状态,其长度总在延长与缩短的变化之中。
3.细胞皮层/肌动蛋白皮层
微丝同微管一样,在体内装配时也有成核作用。所不同的是肌动蛋白纤维的成核作用发生在质膜,因而在很多细胞中肌动蛋白纤维在质膜下的一层由微丝和各种微丝结合蛋白组成的网状结构往往密度较高,称为细胞皮层(cell cortex)或肌动蛋白皮层(actin cortex)。
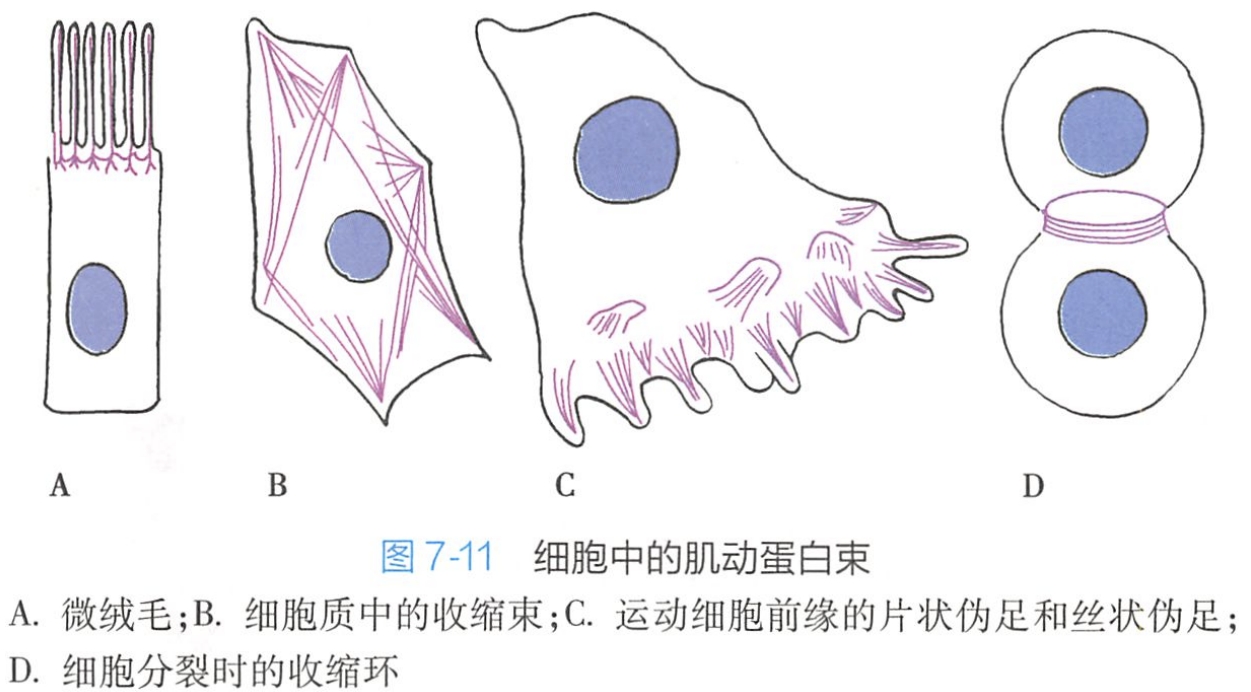
该结构具有很高的动态性,与肌动蛋白一起为细胞膜提供强度和韧性,并维持细胞的形态。如细胞皮层可推动细胞膜形成细长的微刺(microspike),在神经细胞轴突的生长端可形成更长的微穗称丝状伪足(filopodia),还可形成片状伪足(lamellipodia)(图7-l1)。
(三)微丝的组装受多种因素
影响微丝的装配除了受G-肌动蛋白临界浓度的影响,还受ATP、Ca2+、Na+、K+浓度和药物的影响。在含有ADP和Ca2+以及低浓度的Na+、K+的溶液中,微丝趋于解聚而形成肌动蛋白单体;而在含有ATP、 Mg2+和高浓度的Na+、K+的溶液中,肌动蛋白单体则装配成微丝。另外,微丝结合蛋白对微丝的组装也有调控作用。
细胞松弛素B及其衍生物:在细胞内通过与微丝的正端结合起抑制微丝聚合的作用。当将细胞松弛素加到活细胞后,肌动蛋白纤维骨架消失,使动物细胞的各种活动瘫痪,包括细胞的移动、吞噬作用、胞质分裂等。它对微管没有作用,也不抑制肌收缩,因肌纤维中肌动蛋白丝是稳定的结构,不发生聚合与解聚的动态平衡。
鬼笔环肽:同细胞松弛素的作用相反,只与聚合的微丝结合,而不与肌动蛋白单体分子结合。它与聚合的微丝结合后,抑制了微丝的解体,因而破坏了微丝的聚合和解聚的动态平衡。
四、微丝的功能
(一)微丝构成细胞的支架并维持细胞的形态
在细胞中,微丝不能单独发挥作用,必须形成网络结构或束状结构才能发挥作用。细胞的特化结构包括微绒毛(microvilli)和应力纤维(stress fiber)。
1.微绒毛
是质膜顶端表面的指状形突起(见图7-11A)
- 在微绒毛中,由微丝形成的微丝束构成了微绒毛的骨架。
- 另外还有一些微丝结合蛋白,在调节微绒毛长度和保持其形状方面具有重要作用。
- 微绒毛的核心是由20~30个同向平行的微丝组成的束状结构,其中有绒毛蛋白和毛缘蛋白(fimbrin),它们将微丝连接成束,赋予微绒毛结构刚性。
- 另外还有肌球蛋白-Ⅰ(myosin-Ⅰ)和[200 学习/201 细胞生物学/第12章 细胞通讯/第2节 信号转导分子/信号转导分子#(四)钙离子/钙调蛋白信使体系参与细胞收缩、运动、分泌和分裂等重要生命活动|钙调蛋白],它们在微丝束的侧面与微绒毛膜之间形成横桥连接,提供张力以保持微丝束处于微绒毛的中心位置。
微绒毛在小肠吸收上皮细胞最多,使细胞的吸收面积增大20倍,对于增强消化和吸收功能具有重要意义。
2.应力纤维/张力纤维
是真核细胞中广泛存在由微丝束构成的较为稳定的纤维状结构,在细胞内紧邻质膜下方,常与细胞的长轴大致平行并贯穿细胞的全长。这些微丝束具有极性,一端与质膜的特定部位相连,另一端插入到细胞质中,或与中间丝结合。应力纤维具有收缩功能,但不能产生运动,而只能用于维持细胞的形状和赋予细胞韧性和强度。
(二)微丝参与细胞运动
许多动物细胞在进行位置移动时多采用变形运动的方式。如变形虫、巨噬细胞和白细胞以及器官发生时的胚胎细胞等,这些细胞含有丰富的微丝,依赖肌动蛋白和微丝结合蛋白的相互作用,可进行变形运动。
这些细胞通过肌动蛋白聚合使细胞表面形成突起,如片状伪足或丝状伪足。当片状伪足或丝状伪足接触到一片合适的表面时,它们就黏附在上面,这时称为整联蛋白(integrin)的穿膜蛋白就与胞外基质中的分子或与另一细胞表面上的分子结合。同时整联蛋白在胞内面紧紧抓住肌动蛋白丝,从而为细胞内部的肌动蛋白丝系统创造了一个牢靠的锚着点。细胞然后通过内部的收缩产生拉力,利用这一锚着点把自己的身体拉向前。
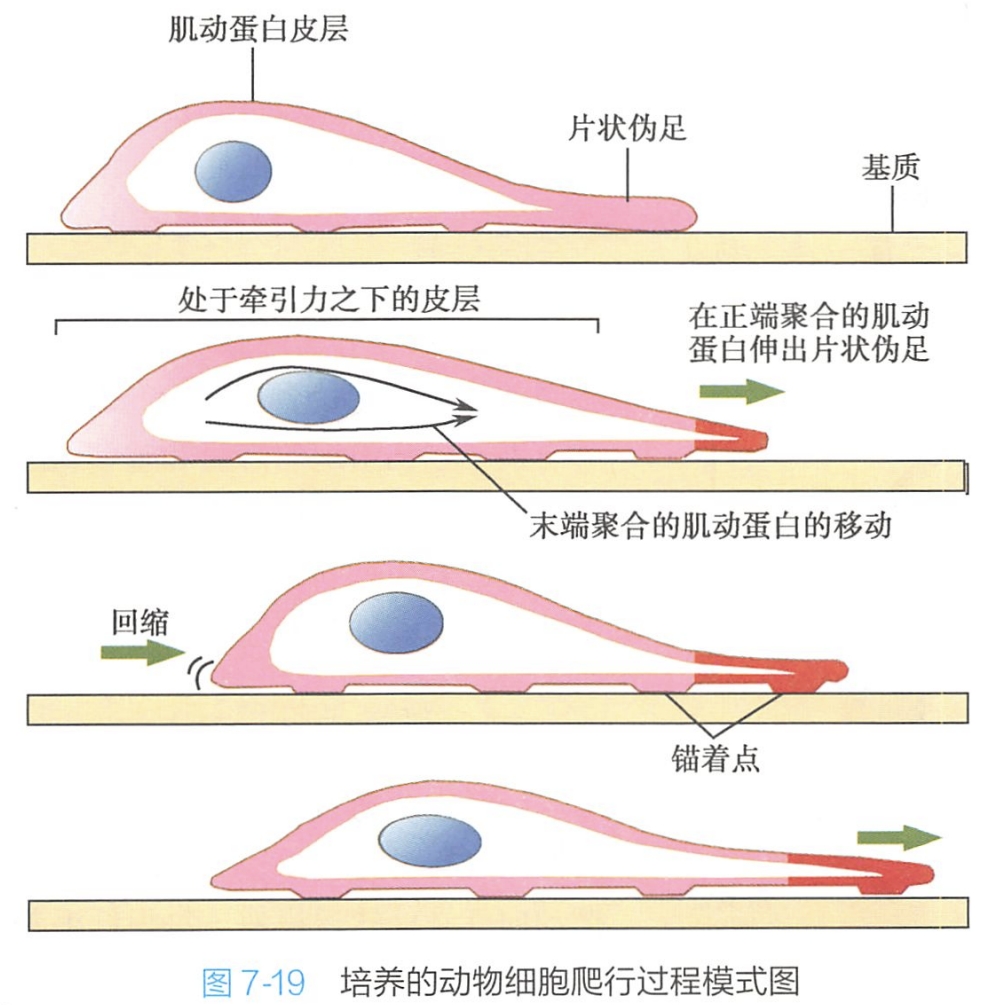
细胞运动是一个高度协同的复杂过程,细胞主要依赖于肌动蛋白和微丝结合蛋白的相互作用进行移动。这种运动可以分为三个过程(图7-19):
1.细胞在它的前端或前沿伸出突起,也叫伪足
- 通过肌动蛋白聚合使细胞表面形成突起。成纤维细胞在爬行过程中,其前缘规律性地伸出薄薄的片状伪足,片状伪足中含有致密的肌动蛋白丝网络,大部分纤维的正端接近细胞质膜。
- 很多细胞也会伸出纤细而坚挺的突起,称为丝状伪足,它们不仅分布在细胞的前缘,而且也分布在细胞表面的其他部分。
2.这些突起附着在其爬行的表面上
当片状伪足或丝状伪足接触到适当的表面时,它们就黏附在上面,这时称为整联蛋白的穿膜蛋白就与胞外基质中的分子或与另一细胞表面上的分子结合,这个运动着的细胞就在基质或另一个细胞上面爬行。同时,在爬行细胞膜的内表面整合蛋白紧紧抓住肌动蛋白丝,从而为爬行细胞内部的肌动蛋白丝系统提供了一个牢固的锚着点。
3.细胞的其余部分通过锚着点上的牵引力将自己向前拉
细胞通过内部的收缩产生拉力,利用锚着点将胞体向前拉动。该过程依赖于肌动蛋白与肌球蛋白的相互作用。但目前尚不清楚这种拉力是如何产生的,是通过收缩胞质中的肌动蛋白丝束呢?还是通过收缩细胞皮层中的肌动蛋白丝网,或是两者兼而有之呢?有待于进一步研究。
(三)微丝参与细胞分裂
动物细胞有丝分裂末期,继核分裂完成后,质膜下皮层由微丝与肌球蛋白-Ⅱ形成的腰带状束,称为收缩环(contractile ring)(见图7-l1D)。收缩环产生的动力将质膜向内拉,细胞的腰部紧缩最终一分为二完成胞质的分裂过程,把两个细胞分隔开。
(四)微丝参与肌肉收缩
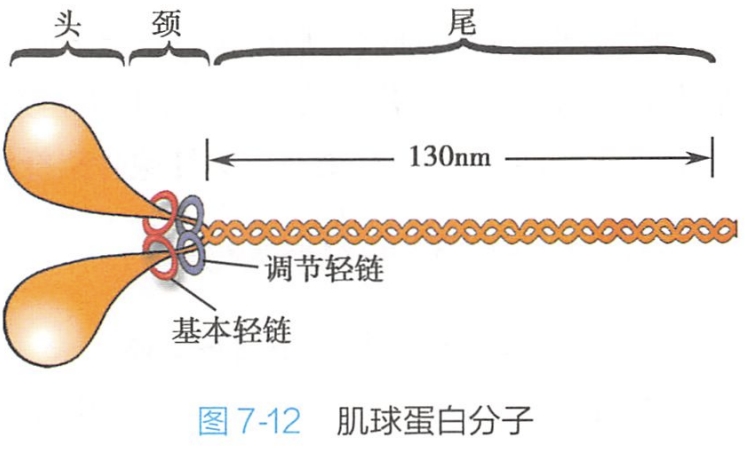
骨骼肌收缩的基本结构单位——肌小节(sar comere)的主要成分是肌原纤维。电镜下观察证明肌原纤维由粗肌丝(thick myofilament)和细肌丝(thin myofilament)组成。粗肌丝由肌球蛋白(myosin)组成(图7-l2)。
细肌丝由肌动蛋白、原肌球蛋白(tropomyosin)和肌钙蛋白(troponin)组成,又称肌动蛋白丝。 #? 肌钙蛋白与钙调蛋白有关系不
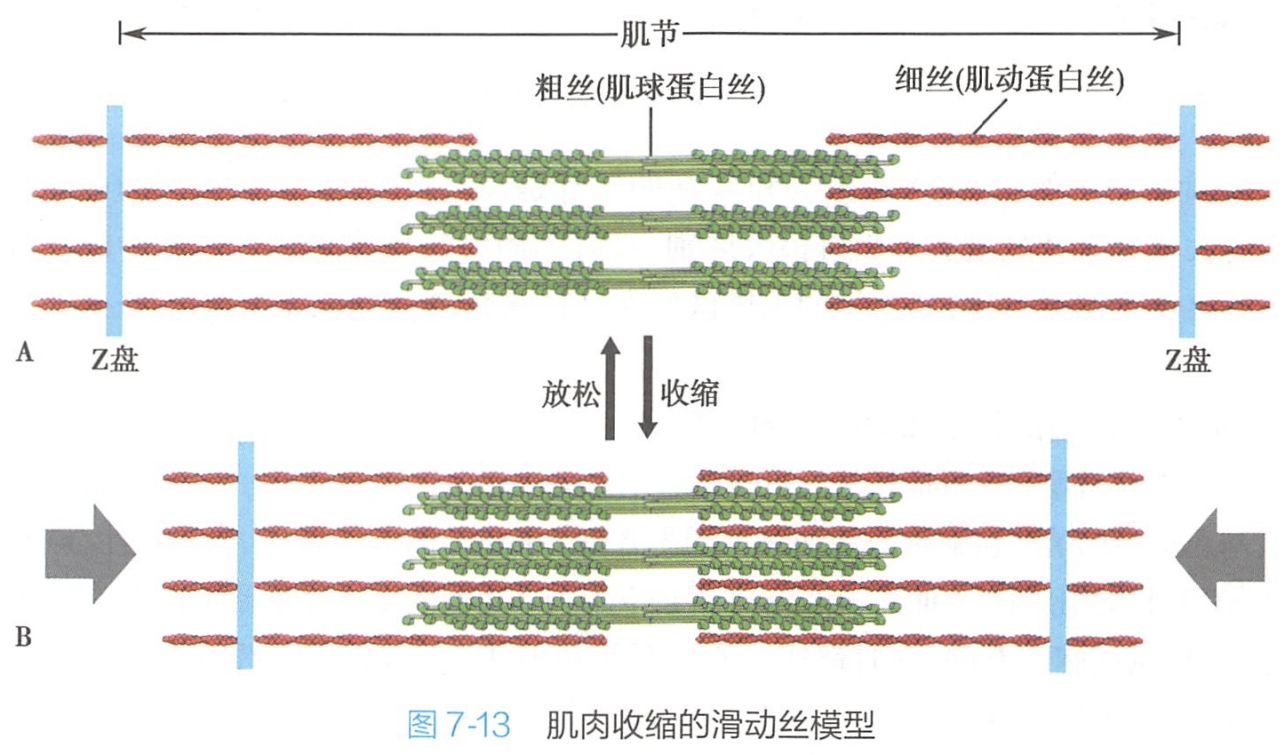
粗肌丝与肌动蛋白细丝相互交错,分布在中线两侧肌细胞的收缩机制可用滑动丝模型(sliding filament model)来解释(图7-l3)。滑动丝模型认为肌细胞收缩是由于粗肌丝与细肌丝之间相互滑动的结果。
肌细胞收缩的分子基础是:肌球蛋白的头部与邻近的细肌丝结合并发生一系列的构象变化,触发肌球蛋白头部沿着细肌丝正端“行走”,从而导致肌肉的收缩。
(五)微丝参与细胞内物质运输
微丝在微丝结合蛋白介导下可与微管一起进行细胞内物质运输,例如小泡的运输,通过肌球蛋白-I与微丝结合,将小泡沿微丝的(-)端向(+)端移动。另外,肌球蛋白-I的尾部与质膜结合,利用其头部可将微丝从一个部位运向另一个部位。
(六)微丝参与细胞内信号传递
细胞表面的受体在受到外界信号作用时,可触发质膜下肌动蛋白的结构变化,从而启动细胞内激酶变化的信号传导过程。